CYTOSKELETON

|
Cells contain elaborate arrays of protein fibers that serve such functions as:
The cytoskeleton is made up of three kinds of protein filaments:
|
Actin Filaments
Monomers of the protein actin polymerize to form long, thin fibers. These are about 8 nm in diameter and, being the thinnest of the cytoskeletal filaments, are also called microfilaments. (In skeletal muscle fibers they are called "thin" filaments.) Some functions of actin filaments:
- form a band just beneath the plasma membrane that
- provides mechanical strength to the cell
- links transmembrane proteins (e.g., cell surface receptors) to cytoplasmic proteins
- pinches dividing animal cells apart during cytokinesis
- generate cytoplasmic streaming in some cells
- generate locomotion in cells such as white blood cells and the amoeba
- interact with myosin ("thick") filaments in skeletal muscle fibers to provide the force of muscular contraction
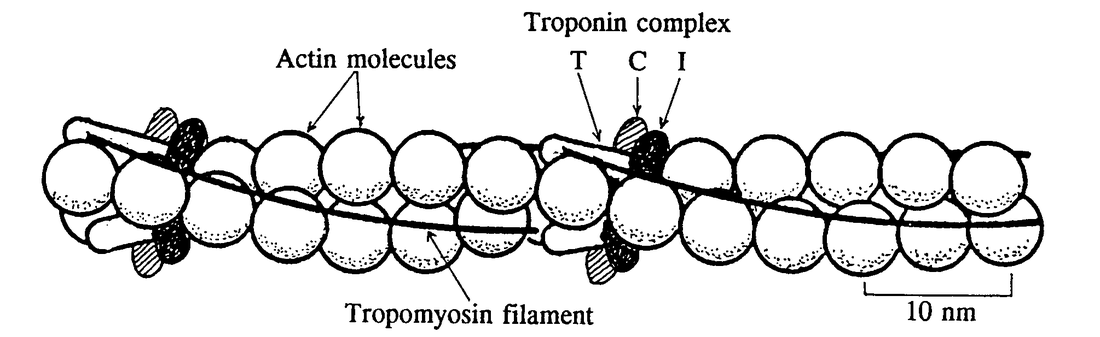
Figure 3.8 Schematic drawing of part of an actin filament showing the relationship between successive pairs of molecules, thin filaments of tropomyosin, and the three types of troponin.
Intermediate Filaments
These cytoplasmic fibers average 10 nm in diameter (and thus are "intermediate" in size between actin filaments (8 nm) and microtubules (25 nm)(as well as of the thick filaments of skeletal muscle fibers).
There are several types of intermediate filament, each constructed from one or more proteins characteristic of it.
Despite their chemical diversity, intermediate filaments play similar roles in the cell: providing a supporting framework within the cell. For example, the nucleus in epithelial cells is held within the cell by a basket like network of intermediate filaments made of keratins. (photo at right)
In the photo (courtesy of Mary Osborn), a fluorescent stain has been used to show the intermediate filaments of keratin in two epithelial cells. Note the basketlike arrangement of filaments around each nucleus.
Different kinds of epithelia use different keratins to build their intermediate filaments. Over 20 different kinds of keratins have been found, although each kind of epithelial cell may use no more than 2 of them. Up to 85% of the dry weight of squamous epithelial cells can consist of keratins.
There are several types of intermediate filament, each constructed from one or more proteins characteristic of it.
- keratins are found in epithelial cells and also form hair and nails;
- nuclear lamins form a meshwork that stabilizes the inner membrane of the nuclear envelope;
- neurofilaments strengthen the long axons of neurons;
- vimentins provide mechanical strength to muscle (and other) cells.
Despite their chemical diversity, intermediate filaments play similar roles in the cell: providing a supporting framework within the cell. For example, the nucleus in epithelial cells is held within the cell by a basket like network of intermediate filaments made of keratins. (photo at right)
In the photo (courtesy of Mary Osborn), a fluorescent stain has been used to show the intermediate filaments of keratin in two epithelial cells. Note the basketlike arrangement of filaments around each nucleus.
Different kinds of epithelia use different keratins to build their intermediate filaments. Over 20 different kinds of keratins have been found, although each kind of epithelial cell may use no more than 2 of them. Up to 85% of the dry weight of squamous epithelial cells can consist of keratins.

Figure 3.9. Intermediate Filament Assembly: Intermediate filaments are built from monomers that associate with each other form dimers. Pairs of dimers then associate in an anti-parallel fashion to form staggered tetramers. Lateral associations between eight tetramers form unit-length filaments, which are able to anneal to each other, end-to-end, to form intermediate filaments
Microtubules
Microtubules
Microtubules
- are straight, hollow cylinders whose wall is made up of a ring of 13 "protofilaments";
- have a diameter of about 25 nm;
- are variable in length but can grow 1000 times as long as they are wide;
- are built by the assembly of dimers of alpha tubulin and beta tubulin;
- are found in both animal and plant cells. In plant cells, microtubules are created at many sites scattered through the cell. In animal cells, the microtubules originate at the centrosome.
- The attached end is called the minus end; the other end is the plus end.
Microtubules
- grow at the plus end by the polymerization of tubulin dimers (powered by the hydrolysis of GTP), and
- shrink by the release of tubulin dimers (depolymerization) at the same end.
Microtubule motors
There are two major groups of microtubule motors:
- kinesins (most of these move toward the plus end of the microtubules) and
- dyneins (which move toward the minus end).
- The rapid transport of organelles, like vesicles and mitochondria, along the axons of neurons takes place along microtubules with their plus ends pointed toward the end of the axon. The motors are kinesins.
- The migration of chromosomes in mitosis and meiosis takes place on microtubules that make up the spindle fibers. Both kinesins and dyneins are used as motors.
- Vincristine, a drug found in the Madagascar periwinkle (a wildflower), binds to tubulin dimers preventing the assembly of microtubules. This halts cells in metaphase of mitosis.
- Taxol®, a drug found in the bark of the Pacific yew, prevents depolymerization of the microtubules of the spindle fiber. This, in turn, stops chromosome movement, and thus prevents the completion of mitosis.

Figure 3.10 Microtubules (MTs) serve as a substrate for kinesin and dynein motor proteins as well as contributing to the shape and division of cells by the interaction with anchoring proteins.
Cilia and Flagella
These whiplike appendages extend from the surface of many types of eukaryotic cells.
StructureBoth cilia and flagella consist of:
Each cilium (and flagellum) grows out from, and remains attached to, a basal body embedded in the cytoplasm. Basal bodies are identical to centrioles and are, in fact, produced by them. For example, one of the centrioles in developing sperm cells — after it has completed its role in the distribution of chromosomes during meiosis — becomes a basal body and produces the flagellum.
- If there are many of them, they are called cilia;
- if only one, or a few, they are flagella. Flagella also tend to be longer than cilia but are otherwise similar in construction.
- For single cells, such as sperm, this enables them to swim.
- For cells anchored in a tissue, like the epithelial cells lining our air passages, this moves liquid over the surface of the cell (e.g., driving particle-laden mucus toward the throat).
StructureBoth cilia and flagella consist of:
- a cylindrical array of 9 filaments consisting of:
- a complete microtubule (the A-microtubule) extending into the tip of the cilium. When a cilium is being disassembled, protein complexes move down from the tip of the cilium traveling along A-microtubules.
- a partial microtubule (the B-microtubule) that doesn't extend as far into the tip. When the cilium is growing, its protein components move up toward the tip of the cilium traveling along B-microtubules.
- cross-bridges of the motor protein dynein that extend from the complete microtubule of one filament to the partial microtubule of the adjacent filament.
- a pair of single microtubules running up through the center of the bundle, producing the "9+2" arrangement.
- The entire assembly is sheathed in a membrane that is an extension of the plasma membrane.
Each cilium (and flagellum) grows out from, and remains attached to, a basal body embedded in the cytoplasm. Basal bodies are identical to centrioles and are, in fact, produced by them. For example, one of the centrioles in developing sperm cells — after it has completed its role in the distribution of chromosomes during meiosis — becomes a basal body and produces the flagellum.

Figure 3.11 Differences Between Cilia and Flagella
